W każdej nauce jest tyle prawdy ile jest w niej matematyki. To przypisywane Kantowi stwierdzenie uzmysławia pewną głęboką prawdę. Nauka o tyle jest nauką, o ile daje jakieś sensowne przewidywania wyników eksperymentów lub obserwacji. A takie przewidywania, które można później potwierdzić, albo odrzucić, są właściwie niemożliwe bez zbudowania jakiegoś modelu danego zjawiska. Modelu matematycznego. Żadna nauka nie jest nauką, o ile nie posiada testowalnych modeli matematycznych. Modele matematyczne wykorzystuje się nawet w takich dziedzinach, których nikt by o podobną perwersję nie podejrzewał, np. w historii.
Można w tym miejscu zwrócić uwagę, że być może niektóre zagadnienia, które pragnęlibyśmy zbadać, jak przykładowo ludzka psychika, są po prostu zbyt skomplikowane, żeby opisujący je choćby z grubsza model matematyczny stworzyć. Być może. Ale jeżeli faktycznie tak jest, to oznacza, że nie da się też ich opisać w żaden inny sposób i zachowanie takich układów staje się całkowicie nieprzewidywalne.
W niniejszym eseju zajmiemy się dziedziną, która, owszem, jest jak najprawdziwszą nauką, czyli posiada stosowne modele matematyczne, ale fakt ten, pomimo szerokiego zainteresowania i kontrowersji, jakie owa dziedzina wzbudza, nie może się jakoś przebić do powszechnej świadomości.
Wyobraźmy sobie, że istnieje zbiór obiektów, które z dostępnych w otaczających ich środowisku zasobów potrafią, na podstawie zakodowanych w sobie instrukcji, budować swoje, niekoniecznie nawet bardzo dokładne kopie, przekazując im ową instrukcję budowania następnych kopii. Po drugie kopie budowane przez jeden określony obiekt, są do niego bardziej podobne, niż kopie budowane przez inne tego rodzaju obiekty, po trzecie, podczas kopiowania instrukcji mogą zachodzić od czasu do czasu błędy, które są oczywiście powielane przez kolejne generacje kopii. Obiekty mające te wymienione właśnie właściwości nazwijmy replikatorami. Czytelnicy zdążyli już się pewnie zorientować, że mowa będzie o organizmach żywych, które są jedynymi znanymi nam obiektami w pełni spełniającymi definicję replikatorów. Dlatego też w dalszej części wywodu, opisując replikatory, będziemy się posługiwać terminologią biologiczną, niemniej warto pamiętać, że pojęcie replikatora jest szersze od pojęcia „organizmu żywego”.
Replikator zatem, buduje swoje kopie, czyli się rozmnaża. Czyni tak na podstawie posiadanej instrukcji, zwanej genotypem. W genotypie zapisane są szczegółowe przepisy na powstanie i wzrost samego replikatora – organizmu, oraz na rodzaj i przebieg replikacji. Sposób realizacji genotypu w świecie fizycznym, czyli kształt, budowa i wygląd replikatora, jego zachowania, zwłaszcza rozrodcze, oraz interakcje, w które replikator wchodzi ze swoim otoczeniem, w tym innymi replikatorami, nazywamy fenotypem. Genotyp podzielony jest na oddzielne instrukcje zwane genami, z których każda odpowiada za jakąś jedną wybraną cechę fenotypu. Istnieją także geny, w biologii zresztą wcale za geny sensu stricte nie uważane, które żadnych cech fenotypowych nie kodują. Oczywiście instrukcja genetyczna nie jest jedynym czynnikiem kształtującym fenotyp. Nie mniej bowiem istotną rolę odgrywają w tej materii czynniki środowiskowe. Jednak tylko te cechy fenotypu, które zakodowane są w genotypie dziedziczone są przez kolejne pokolenia replikatorów. Cechy zaś nabyte w trakcie życia replikatora, wbrew twierdzeniom lamarkistów i ich komunistycznej proweniencji ideologicznych patronów, dziedziczone nie są. Zachodzące w dłuższym okresie czasowym zmiany w dziedzicznych cechach fenotypu, a co za tym idzie zmiany w genotypie, nazywa się ewolucją. Ewolucja w tym kontekście, to nie jakakolwiek zmiana, ale przekazywana z pokolenia na pokolenia zmiana w rozkładzie cech fenotypowych, czyli naprawdę w rozkładzie programujących je genów. W tym sensie ewoluują replikatory, oraz np. ludzkie języki, które przynajmniej niektóre cechy replikatorów posiadają. Natomiast gwiazdy, planety i sam Wszechświat, chociaż zmieniają się w czasie, i czasami pisze się o ich „ewolucji” to jednak nie jest to prawdziwa ewolucja.
W niniejszym eseju autor podejmie próbę skonstruowania, odpowiedniego modelu matematycznego opisującego tak rozumianą ewolucję. Postaramy się zatem stworzyć teorię ewolucji. Oczywiście w nauce, taka teoria, zwana od nazwiska naukowca, który jako pierwszy sformułował część jej założeń, darwinizmem, od dawna istnieje i funkcjonuje, ale z trudnych do zrozumienia powodów, jej istnienia nie chcą zauważać zarówno ci, którzy „darwinizm” (czyli tak naprawdę to co oni sami uważają za „darwinizm”) w sposób iście religijny czczą i wysławiają, jak i ci, którzy ów mityczny „darwinizm” zwalczają.
Zaznaczyć trzeba, że teoria ewolucji opisuje zjawisko ewolucji, czyli zgodnie z podaną uprzednio definicją zmianę rozkładu dziedzicznych (genotypowych) cech replikatorów zachodzącą w czasie. I tyle. Nie zajmuje się teoria ewolucji dochodzeniem jak powstały pierwsze replikatory, nie zajmuje się budową i pochodzeniem Ziemi, Układu Słonecznego, i całego Wszechświata. Nie odpowie nam też na pytanie, dlaczego Wszechświat ma takie właściwości, jakie ma, dlaczego prawa fizyki w nim obowiązujące są takie a nie inne, chyba, że uda się komuś wykazać, a takie próby są jak najbardziej serio podejmowane, że Wszechświat również jest replikatorem.
Przyjrzyjmy się teraz genom. Nie będziemy wnikać, w jakiej fizycznej postaci istnieją geny w organizmie, w jaki sposób instrukcje genetyczne są przekładane na cechy fenotypowe, czy jak konkretnie geny się kopiują przy rozmnażaniu – replikacji. Wystarczy nam, że określone geny odpowiadają za określone cechy fenotypowe i że w trakcie rozmnażania są owe geny powielane i przekazywane kolejnemu pokoleniu replikatorów. Ponieważ jednak nic na tym świecie nie jest doskonałe, w trakcie owego powielania i przekazywania genetycznej instrukcji zdarzają się błędy. Błędy takie w biologii są znane jako mutacje i istnieje w nauce pogląd, zwany mutacjonizmem, który właśnie mutacjom przypisuje rolę decydującego mechanizmu napędzającego ewolucję. Istnieje wiele różnych rodzajów mutacji, ale nam wystarczy założenie, że mutacja zmienia wersję a danego genu, na wersję A. (a=>A) Prawdopodobieństwo zajścia takiej mutacji wynosi PA. Jeżeli w danej populacji o łącznej liczebności danych genów, co w przypadku organizmów rozmnażających się bezpłciowo odpowiada liczebności osobników, N istnieje już Na wersji genu a i NA wersji genu A czyli zachodzi równość (Na+NA=N), wtedy zmiana proporcji występowania tych dwóch wersji genu wynosiłaby
dNA = (N-NA)*PA
Oczywiście naiwnością byłoby oczekiwanie, że przypadkowe mutacje mogą działać tylko w jedną stronę. Gdyby tak było, to wcześniej, czy później gen a, zostałby wyparty przez wersję A całkowicie, tylko z powodu istnienia możliwości mutacji. Byłby to mutacjonizm posunięty do skrajności. Najczęściej bywa tak, że możliwe są również mutacje wsteczne, zamieniające gen A z powrotem w jego wersję a. Niech prawdopodobieństwo takiej wstecznej mutacji wynosi Pa. Wtedy nasze równanie mutacjonistyczne będzie wyglądać tak:
dNA = (N-NA)*PA-NA*Pa
Po podzieleniu stronami przez liczbę genów N otrzymamy względną częstość genu A Q, czyli ostateczną wersję równania mutacjonistycznego.
(1) dQ/dt = (1-Q)*PA-Q*Pa
Równanie to można rozwiązać albo numerycznie, poprzez podstawianie i iterację kolejnych wartości Q, albo wprost analitycznie. Zastosujemy ten drugi sposób, bo w miarę rozbudowy naszej teorii ewolucji, utracimy w znacznej mierze ten luksus i zostaną nam tylko iteracje. Załóżmy rozsądnie, że prawdopodobieństwo zajścia mutacji w obie strony jest jednakowe (PA = Pa = P). Po odpowiednich podstawieniach, przekształceniach i całkowaniu otrzymujemy wzór.
(2) Q(t) = 0,5*((2*Q0-1)*exp(-2*P*t)+1)
Gdzie Q0 to początkowa częstość genu A. Jeżeli Q0 = 0, to równanie (2) zredukuje się nam do postaci
(3) Q(t) = 0,5*(1-exp(-2*P*t))
Z równań tych wynika, że mutująca populacja dąży do pewnego stanu wysycenia, w której mutacje zachodzące w obie strony się równoważą. Możemy znaleźć ową równowagową częstość zmutowanego genu Q, przyrównując równanie (1) do zera, co oznacza brak zmian Q. Otrzymamy wtedy
Qr = PA/(Pa+PA)
A przekształcając równanie (3) możemy określić czas t pokoleń, po jakim częstość zmutowanego genu, startując od zera, osiągnie określoną wartość Q.
T= -ln(1-2*Q)/(2*P)
Zauważmy, że zarówno Q równowagi, jak i czas dojścia do określonej częstości zależy wyłącznie od prawdopodobieństwa zajścia mutacji P. Tak jest oczywiście średnio. A średnia ma to do siebie, że możliwe są od niej odchyłki. Ponieważ mutacje są zdarzeniami losowymi, takie odchyłki od średniej będą się oczywiście zdarzać. Miarą takiego rozrzutu jest wielkość zwana odchyleniem standardowym. Nie wdając się w detale, jest ono proporcjonalne do pierwiastka kwadratowego z wielkości populacji N, czyli wraz ze wzrostem liczebności populacji rośnie wolniej niż sama populacja. Im populacja większa, tym względna odchyłka od wartości średniej będzie mniejsza. Na poniższym wykresie, zaprezentowano wzrost częstości genu pod wpływem mutacji, czyli mutacjonizm. Zarówno wartość średnią, jak i realną z uwzględnieniem istnienia przypadkowych odchyłek od średniej, przy założeniu, że wielkość populacji wynosi 1000 kopi genu. Czytelnik sam zechce obliczyć, jakie przyjęto prawdopodobieństwo wystąpienia mutacji P.
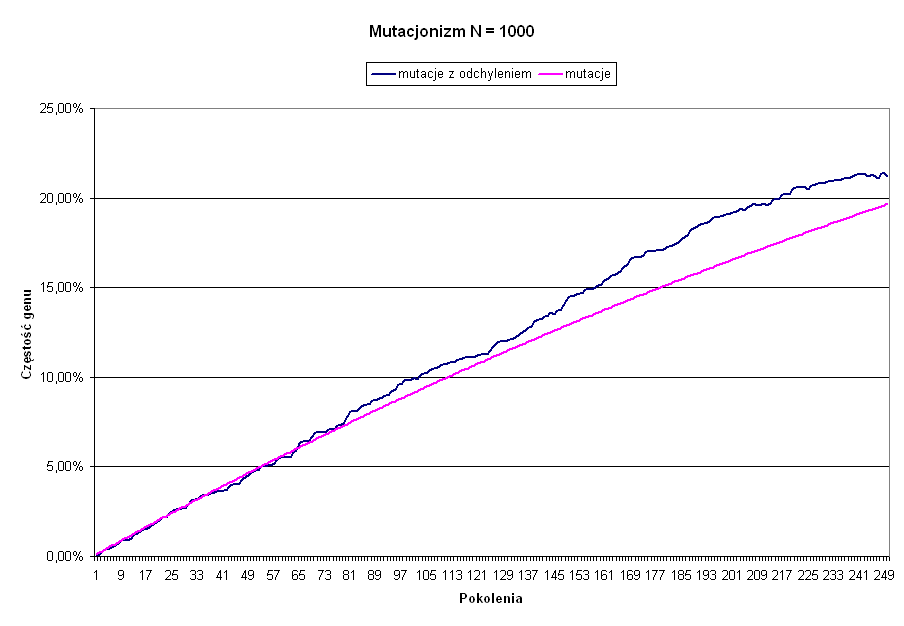
Oczywiście opisane przed chwilą przypadkowe odchylenia od średniej nie dotyczą tylko mutacji. Jeżeli w populacji istnieje już zmienność genetyczna, czyli istnieją różne warianty tego samego genu, to należy oczekiwać, że częstość tych wariantów w populacji nie pozostanie stała. W każdym następnym pokoleniu może się ona losowo zmienić w dowolną stronę i nie ma żadnej gwarancji, że w kolejnym ruchu powróci do stanu poprzedniego. Można to porównać do losowego rzutu monetą, w którym prawdopodobieństwo wypadnięcia orła, lub reszki nie zależy od tego, jaki był wynik poprzedniego rzutu. Po pewnym więc czasie, częstość danego genu w populacji może przyjąć każdą wartość, z 0, czyli całkowitym wyginięciem, oraz 100 %, czyli z wyparciem z populacji wszystkich innych wariantów danego genu, włącznie. Zjawisko to nazywa się dryfem genetycznym i istnieje w nauce pogląd, zwany neutralizmem, który właśnie dryfowi genetycznemu przypisuje rolę decydującego mechanizmu napędzającego ewolucję. Podobnie jak w przypadku mutacji, losowe odchyłki od istniejącej częstości genu są proporcjonalne do pierwiastka kwadratowego z wielkości populacji, i maleją wraz ze wzrostem liczebności populacji. Na poniższym wykresie zaprezentowano przebieg ewolucji neutralnej, dla pierwotnej częstości równej 50% i różnych wielkości populacji.
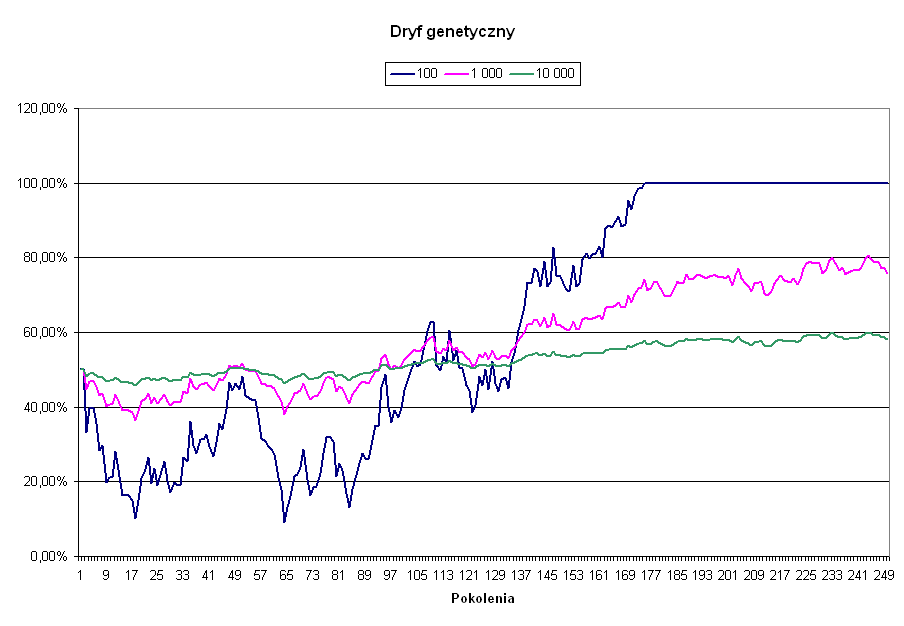
Jak widać, im mniejsza populacja, tym szybciej dany gen zostanie utrwalony, bądź z niej usunięty. Jeżeli istnieje jakieś niezerowe prawdopodobieństwo mutacji, to wcześniej czy później, w każdej skończonej populacji, mutant ten wyprze poprzednie wersje genu. Ewolucja pod wpływem czystego przypadku jak najbardziej zatem zachodzi, chociaż tempo jej dla większych populacji jest bardzo powolne.
Omówiliśmy zatem dwa człony budowanej przez nas teorii ewolucji. Mutacjonizm i neutralizm. Oba opierają się na przypadku. Przypadkowych mutacjach i przypadkowym dryfie genetycznym. Niemniej, wbrew dominującej w niektórych środowiskach opinii, przypadek to nie to samo, co chaos. Zdarzeniami losowymi rządzą ścisłe matematyczne reguły i dlatego możliwe jest, co właśnie uczyniliśmy, zbudowanie modelu matematycznego opisującego czysto przypadkową ewolucję. Ale przecież ewolucja to nie tylko przypadek. Gdyby tak było, to nie wiedzący nic o mutacjonizmie i neutralizmie Karol Darwin, nigdy by nie wstrząsnął nauką i społeczeństwem. Odkrył on jednak istnienie siły ewolucyjnej znacznie od neutralizmu i mutacjonizmu potężniejszej, a przy tym nie mającej nic wspólnego z losowością i przypadkiem. Tą właśnie siłą zajmiemy się na kolejnym etapie budowy naszej teorii ewolucji.
PS W przypływie niewiarygodnej wręcz łaskawości i miłości bliźniego, autor zaproponował publikację powyższego tekstu portalowi (ir)racjonalista.pl Redakcja portalu szansę na skokowe podniesienie swojego poziomu merytorycznego odrzuciła w tzw „pierwszym czytaniu”, nie proponując nawet żadnych zmian, nie odsyłając do recenzentów, etc. Jest to juz drugi taki przypadek odrzucenia merytorycznego tekstu z tego zakresu.
Merytoryczne prace w zakresie teorii ewolucji nie znajdują się w kręgu tamtejszych zainteresowań.

z tą mutacją wsteczną to lekkie uproszczenie. Owszem istnieje taki termin w biologii ale chodzi w nim zgoła o coś innego, poza tym dotyczy pojedynczego genu .Zaś powrót do pierwotnego układ nukleotydów nie jest też dziełem przypadku. Przyczyny są złożone ale nie do końca wyjaśnione, więc meritum w tej kwestii to raczej marna spekulacja ubrana w płaszczyk pozornej wiedzy. Równie warta jest teoria o siłach witalnych, entelechiach czy arystotelesowska przyczyna celowa. W Twoim rozumieniu to tak jakbyśmy chcieli założyć istnienie prawdopodobieństwa: iż spontanicznie rozsypując worek piasku a następnie z powrotem pakując piach do wora uzyskamy identyczne przestrzenne sąsiedztwo /relacje każdego ziarna z osobna ze stanu przed rozsypaniem. Zastanawiam się co chcesz w ogóle udowodnić? Że istnieje ewolucja? To chyba truizm nawet dla kreacjonistów 🙂 Współczesny spór toczy się o jej faktyczny przebieg i charakter a nie o to czy istnieje. Te wzory, którymi się posiłkujesz to nie dowód ani definicja ewolucji tylko przykład jak funkcjonuje prawdopodobieństwo; co wyniknie przy losowaniu kulek z jakiegoś zbioru etc. O ewolucji nie ma tu mowy. Nota bene sama ewolucja nie jest wynalazkiem Darwina. Więcej u niego wartości literackich niźli naukowych – oczywiście w rozumieniu kantowskim 😉
PolubieniePolubienie
Niestety Lyman nic nie zrozumiał z tego co napisałem. Jezeli istnieją możliwe dwa stany genu (A i a), to istnieją równiez i prawdopodobieństwa przejścia A w a i a w A. Prawdopodobieństwa te nie muszą być równe. I jużCałe to gadanie o workach piasku to tylko gadanie.A kreacjoniści, owszem, głoszą, że ewolucja nie zachodzi. Że powstanie nowego gatunku z dotychczas istniejącego, jest niemozliwe
PolubieniePolubienie
ależ Lyman zrozumiał.Lyman tylko zauważa,że wyniki z takiej nauki nie powiedzą nam nic o ewolucji (czy PRAWDOPODOBNYM procesie ewolucji(np w czasie) a jedynie o prawdopodobieństwie wystąpienia zmian jakichś elementów i to w warunkach idealnych. Nie zarzucam błędów w rachunkach. Jest to bardzo dobry przykład ćwiczenia matematycznego dla licealistów. Mamy czytelne zbiory, elementy etc.. Ten model jednak jest za prymitywny aby cokolwiek określić w ewolucji biologicznej. Wystarczający jednak by słupkami i statystykami mamić jakiś odsetek społeczny bo przecież nie masy. Dlatego zastanawiam się do kogo to ma trafić? Nie do brukowców w dziale sensacje(bo za trudne) ..ale do prawdziwych naukowców chyba też nie. Rozumiem,że do ćwierćinteligentów lub właśnie kreacjonistów.Ok. Może ich przekonasz. Przypomina mi to trochę Dawkinsa, który podobnymi argumentami „naukowymi” próbuje dowieść masom społecznym,że nie istnieje bóg. Choć Ty optujesz raczej w kierunku Laplace’a,który zakładał,że przez odpowiedni wzór matem. możemy przewidzieć każde zdarzenie i całą przyszłość. Ale to już determinizm do którego się chyba nie przychylasz 😉
PolubieniePolubienie
Niestety jednak Lyman nie zrozumiał :)Proces ewolucyjny, taki jak jest obserwowany w skali makro, składa się właśnie z takich małych, drobnych fragmencików. W eseju pokazałem jak nowe cechy w populacji powstają i jak się w niej przypadkowo rozprzestrzeniają. Takich cech oczywiscie może być bardzo dużo, ale kazda zachowuje sie w ten właśnie opisany przez pilastra sposób.Oczywiscie wyłącznie wtedy, kiedy ta cecha jest neutralna, czyli nie wpływa na proces replikacji. Jezeli jednak wpływa, to wtedy mamy do czynienia nie z dryfem, ale z doborem naturalnym, o czym będzie mowa w następnym odcinku.Krytykowanie tego eseju pod pretekstem, że opisuje tylko pewien wyidealizowany fragment ewolucji, przypomina krytykowanie mechaniki bryły sztywnej, ze zajmuje się tylko „idealnymi”, „platońskimi” walcami, kulami, etc i w zwiazku z tym nie jest nam w stanie NIC powiedzeić o prawdizwych fizycznie istniejących bryłach.
PolubieniePolubienie
Witaj,chłopie – ale żeś się zapalił do tej „religii”, oh, pardon – „nauki”! :-)Powiem tak: proszę, podaj mi przykład pozytywnej mutacji z przyrody.Chciałbym również się dowiedzieć czy wszystkie zwierzęta (w tym ludzie) oraz rośliny są ze sobą spokrewione, bo tak pisał Darwin.Jak sądzisz, co wiesz?Czeka na odpowiedź o pozdrawiaJacek W.jacklondon@hotmail.co.uk
PolubieniePolubienie
> Powiem tak: proszę, podaj mi przykład pozytywnej mutacji z> przyrody.Co Jack rozumie przez „pozytywną mutację”?> Chciałbym również się dowiedzieć czy wszystkie zwierzęta> (w tym ludzie) oraz rośliny są ze sobą spokrewione, Eukarionty? Owszem na pewno tak. Co do prokariontów prawie na pewno tak (ostatnio pojawiły się doniesienia o alterenatywnym „życiu arsenowym” na poziomie mikroorganizmów)> bo tak> pisał Darwin.Nie tylko dlatego 🙂
PolubieniePolubienie
Małe przegięcie z tym co piszesz i niezrozumienie tematu. A jeżeli hccesz przesledzić matematyczne modele ewolucji to prosze bardzo. http://www.youtube.com/watch?v=9G-bxGenkoMhttp://www.youtube.com/watch?v=n_EPvkNnLR0&feature=relatedTroche uprościłeś sprawe nawet na polu matematyki. A portal racjonalista.pl jest zlewicowany, oczywiscie twój tekst powinien się pojawić by mozna go było zmasakrowac :-).
PolubieniePolubienie
> Małe przegięcie z tym co piszesz i niezrozumienie tematu.Na czym polega niezrozumienie tematu?> Troche uprościłeś sprawe nawet na polu matematyki.Na czym polegają te uproszczenia?
PolubieniePolubienie